

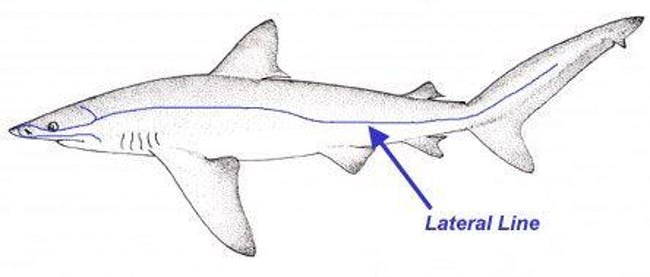
The Lateral Line is one of the most unique sense organs in the animal kingdom. Possessed by most fish, the lateral line enables fish to sense vibrations and movement in the water around them. This article will discuss how the lateral line works and its implications in the origins debate.
The lateral line is not a single organ, but rather many. Just beneath the skin is a line of specialized mechanoreceptors called neuromasts. A neuromast comes in two varieties. The first is known as the superficial neuromast because it is found on the surface of the fishes scales. The second variety is known as the canal neuromast. They are found beneath the scales in tiny liquid-filled canals. Both types are made of a special receptor called hair cells. These hair cells are just above the scales and detect the movement of water across the fishes body. Water is a conductor of sound, energy, and motion. The hair cells detect this sound, energy, and motion and pass it along to specialized nerve cells at the base of the hair cells. This information is then passed to the brain, which uses it to form a mental image of the fishes surroundings.
Fish rely heavily on their lateral lines, more so even than some of their other senses. For example, it has been demonstrated that blind fish can swim in a school of fish and have no collisions or other such difficulties. Fish which are not blind, but have had their lateral line disabled cannot do to. The ability to see an outline picture, even without eyes, is immensely useful. The lateral line enables fish to navigate in pitch darkness, relying on water currents and small recoils from obstacles. Fish also can use the lateral line to orient themselves in the water or hunt down escaping prey. The ripples which are given off by a rapidly swimming fish roll over the predators lateral line, enabling it to form a picture of its prey’s path, even as the prey attempts to flee.
Sharks and rays take this functionality several steps further. They have specialized cells in the snout known as the Ampullae of Lorenzini. These function as electroreceptors, enabling these predators to sense tiny electrical pulses in the water. These special receptors are found on the surface of the skin as tiny pores, mostly on the underside of the shark’s jaw. These pores lead back to a gelatinous chamber where the electrical charges detected are passed on to nerves which carry them to the brain. The brain can form an image based on these signals that will indicate where the signals are coming from. The signals themselves are generated by the movement of fish muscles as they swim. Sharks often have thousands of the Ampullae of Lorenzini, usually concentrated in the nose and under the jaw. This enables them to track their prey through water in pitch darkness or murk.
The lateral line and the Ampullae of Lorenzini have massive implications in the origins debate. They are separate structures so they will be examined separately. The lateral line is found in fish and has been reported in some amphibians. However, its existence is not in question. What is in question is where it came from. For example, why would fish evolve modified hair cells when fish have no hair, to begin with? Where did the gene for the hair come from? Hair itself does not appear until the first mammals in the evolutionary tree so how could modified hair cells appear possibly hundreds of millions of years earlier? Why would a fish evolve a lateral line in the first place? If the nerve endings were present, but not the line itself, the endings would be a waste of energy. If the line and its specialized hair cells were present but not the nerve endings, there would be no way to get the information to the brain where it could be interpreted. The whole thing had to be in place, from the beginning, or none of it would work. However, the lateral line does not come close to the challenge posed by the Amullae of Lorenzini.
The Ampullae of Lorenzini is a massive challenge to evolutionary precepts. They are unique to fish. They are also uniquely designed to detect electrical impulses. The gel found in the canals has the highest conductivity of any biological material. This is a little too coincidental to be a result of blind random chance. In order for the Ampullae of Lorenzini to work, the canals, nervous systems, and gel, all have to be present. Moreover, the gel has to have the high conductivity that it has so that sharks and rays can detect the electrical signals given off by muscular movement of their prey. If any of these conditions are not met, the ampullae will not function. Therefore, all conditions must be met at the beginning. The ampullae could not have evolved gradually, as it would not have worked for the vast majority of its development. Evolution would have weeded out fish wasting energy on parts that were non-functional. However, the complexity of the ampullae goes beyond that. Assuming that the four parts could somehow evolve, a mechanism for this evolution must be found. Why would sharks, king predators, even need the ampullae in the first place? How could such a complex system arise by random chance? This question must be answered. If it cannot be, evolution is left with the problem of how the ampullae developed. It then must stare down the barrel of admitting the need for design or reconstructing the entire theory.
Evolution has to answer how these two incredibly designed organs came into being. If the theory cannot do so, then it fails as a theory. Evolutionists can point to supposed common ancestors and missing links all they want, but until they have the ability explain how every single system, every organ, every cell type, came into existence, their theory is not proved.
